反刍动物头骨的多样化:从微观进化过程到宏观进化模式
进化生物学家的目标是基于人口水平变异的比较数据集,在微观进化过程和宏观进化模式之间形成基本联系。在一份关于科学进展的新报告中,芝加哥大学和美国杰克逊实验室的Daniel R. Rhoda和一组进化生物学科学家分析了之前发表的反刍动物(哺乳动物)颅骨数据集。
结果通过高度保守的颅面进化同种异素测量(CREA)存在偏差,其中较大的物种表现出成比例的更长的脸。结果强调该特征是阻力最小的进化线,促进了与浏览器-食草动物谱系一致的形态多样性。结果表明,种群水平的约束如何在宏观进化尺度上产生高度定向的表型进化模式。这项工作阐明了探索颅面进化同种异体测量在哺乳动物分支中的作用。
颅面进化同种异体测量 (CEA)
自然选择影响种群中的表型变异,其中种群的发展对选择做出反应。最大变异量的方向称为最小阻力线(LLR),代表最大进化变化的潜在方向。如果选择与阻力最小的线保持一致,生物学家预计种群将以直接路径进化到适应性峰值。但是,如果选择朝向其他地方,则对选择的响应将重新调整为阻力最小的线。因此,适应性景观与种群水平上物种内变异限制之间的相互作用决定了表型进化的路径。
进化生物学家试图将生物多样性的全球模式解释为生物学研究的基本目标,以揭示宏观进化模式背后的微观进化机制。他们预计这些模式会影响人口层面对宏观进化的限制。在这项工作中,Rhoda等人提出了强有力的证据,证明颅面进化同种异构(即生物随大小变化的特征)对微观和宏观进化的影响,可以沿着最小阻力线进行探索,以促进形态多样性。
哺乳动物头骨的高度保守图案
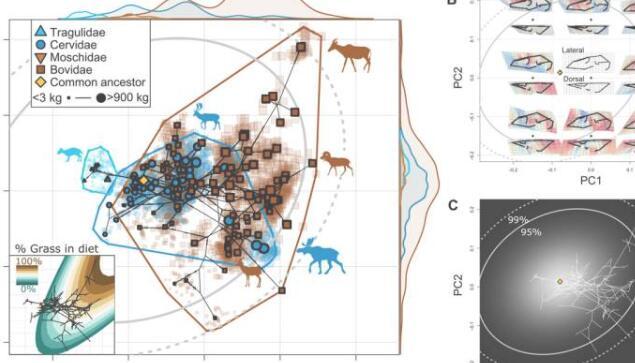
哺乳动物头骨保持着高度保守的个体发育和进化同种异构模式,其中较大的物种具有成比例的更长的脸,作为颅面进化同种异构的一个例子。虽然同种异体是表型进化的一个限制,但它为极端表型的出现提供了机会,而没有发育新颖性。
例如,较大的短脸哺乳动物是不可能的,而长脸,遥不可及的表型是由于这种同种异体测量而出现的。因此,颅面进化同种异体测量法作为阻力最小的进化线出现,简单地表明由于头骨形成的大小相关限制,较大的哺乳动物具有更长的脸。古生物学家注意到物种之间的大小和头骨形状之间存在很强的关系,其中较大的物种代表更长的脸,并且进化同种异体的斜率在亚科之间显着变化。
物种之间的颅骨形状
示例工作显示,由于鼻缩幅度较大,Dik-dik的颅骨形状与其他小型反刍动物不同。然而,所有小型反刍动物的脸都比它们更大、更长的亲戚短。使用系统发育主成分分析,该团队显示出明显的大小趋势主导了变异的主轴,以进一步加强结果。Rhoda等人重建了进化枝之间的种间形态空间,以更好地了解不同的进化模式,同时探索颅面进化同种异构。
研究小组注意到并证实了分析中的形态空间相似性是由于同种异构 - 对牛头骨多样化的主要影响。研究人员注意到牛颅骨进化变异的明显大小趋势,其中大型放牧部落Alcephini和小型浏览部落Neotragini相对于其观察到的进化速度的多样化程度超过预期。这项工作与牛和子宫颈多样化的宏观进化模式一致,这些模式似乎与它们的祖先相去甚远。这些数据代表了新近纪时期反刍动物饮食的进化不稳定性,同时说明了整个新生代Perissodactlys的多样性下降。
CREA跨物种的适应性意义和微观进化的动力学
Rhoda等人研究了与浏览器-食草动物连续体相关的形状变化,并不清楚大型放牧物种(如Alcelaphini)的超长面颅骨是否是由于(1)直接选择更适合放牧的颅骨,巧合的是长面,或者(2)与觅食生态学的选择无关。在后一种情况下,他们的假设是,在与放牧相关的体型增加过程中,更长的脸被动进化,由于食物摄入和加工之间的密切联系,这似乎不太可能。
在任何一种可能的情况下,觅食生态学都为大小的进化变化提供了适应性价值,并影响了反刍动物颅骨形态的多样化。Rhoda等人研究了颅面进化同种异体测量(CREA)是否是反刍动物节肢动物中抵抗力最小的进化系,并分析了其对形态多样性的影响。他们直接测量了与假设的CREA相关的变异轴,以询问其与种群水平上进化变异的关系,并进行系统发育广义最小二乘回归,以测量形态发散与相对于CREA的发散角之间的关联。
展望
通过这种方式,Daniel R. Rhoda及其同事研究了哺乳动物(反刍动物)头骨多样化过程中宏观进化模式背后的微观进化过程。他们探索了颅面进化同种异体测量法(CREA)作为最小阻力线(LLR)的一个方面,因此当他们在工作期间生成物种矩阵时,结果与CREA密切相关;表明该物种与祖先的分歧比那些不太对齐的物种更远。
这项工作研究了进化枝之间差异的内在和外在因素,以及它们相对于浏览器-食草动物谱系中牛和子宫颈的多样化模式。并强调了内在因素对反刍动物颅骨形态多样化的影响。这些结果广泛证明了内在限制对宏观进化模式的影响,使科学家们能够设想进一步研究CREA在其他哺乳动物分支中的作用。
免责声明:本文由用户上传,与本网站立场无关。财经信息仅供读者参考,并不构成投资建议。投资者据此操作,风险自担。 如有侵权请联系删除!
-
安徽淮南地区的长安汽车经销商近期对2025款启源Q05车型的市场策略进行了调整,以进一步吸引消费者。作为长安汽...浏览全文>>
-
近年来,豪华SUV市场持续升温,而作为大众旗下的高端品牌,途锐凭借其卓越的性能与豪华配置,一直深受消费者的...浏览全文>>
-
在新能源汽车市场中,一汽-大众ID 4 CROZZ凭借其出色的性价比和丰富的配置吸引了众多消费者的关注。作为一款...浏览全文>>
-
近年来,随着国内汽车市场的快速发展,越来越多的消费者开始关注性价比高的小型车。QQ多米作为一款备受关注的...浏览全文>>
-
在当今的汽车市场中,选择一款高性价比的车型是许多消费者的重要考量。对于追求品质与经济平衡的购车者来说,...浏览全文>>
-
阜阳地区消费者对上汽大众途岳的关注度一直很高,尤其是2025款途岳的上市更是引发了广泛关注。作为一款紧凑型S...浏览全文>>
-
天津滨海长安猎手K50作为一款备受关注的车型,在市场上拥有较高的关注度。这款车型以其出色的性能和合理的价格...浏览全文>>
-
近年来,随着新能源汽车的普及,插电混动车型因其兼顾燃油经济性和驾驶性能的特点,受到了越来越多消费者的青...浏览全文>>
-
途观X作为上汽大众旗下的高端轿跑SUV车型,凭借其时尚动感的外观设计和丰富的科技配置,在市场上一直备受关注...浏览全文>>
-
在选择SUV车型时,上汽大众途昂凭借其宽敞的空间和强大的性能成为不少消费者的首选。对于蚌埠地区的消费者来说...浏览全文>>
- 天津滨海长安猎手K50多少钱 2025款落地价,最低售价18.29万起,赶紧行动
- 天津滨海ID.4 CROZZ 2025新款价格,买车省钱秘籍
- 安徽淮南途昂X多少钱?性价比超高的选车秘籍
- 福特领睿试驾,开启完美驾驭之旅
- 比亚迪海豹05DM-i试驾预约流程
- 凯迪拉克CT5预约试驾,从预约到试驾的完美旅程
- 滁州途观X落地价,各配置车型售价全知晓
- 极狐 阿尔法T6试驾,开启完美驾驭之旅
- 长安欧尚520试驾操作指南
- QQ多米试驾需要注意什么
- 长安Lumin试驾,感受豪华与科技的完美融合
- 安庆长安猎手K50最新价格2025款与配置的完美平衡
- 池州迈腾GTE最新价格2022款全解,买车必看的省钱秘籍
- 安徽亳州ID.6 X多少钱?购车全攻略来袭
- 五菱雪宝试驾,开启豪华驾驶之旅
- 马自达EZ-6预约试驾,新手必看的详细流程
- 安徽亳州T-ROC探歌落地价实惠,配置丰富,不容错过
- 滁州高尔夫GTI价格大揭秘,买车前必看
- 东莞威然落地价,最低售价22.99万起优惠不等人
- 试驾王牌M7,体验豪华驾乘的乐趣